
文責・島田草太朗
複雑な生態系を少しでも理解し、さまざまな自然を徹底的に究明したい京大!バイオスクープ。今回のご依頼はこちら「カタツムリはどうやって歩いているの?」
陸貝
陸生の軟体動物(貝類;Molluscs)のこと。(今の所)陸貝は腹足類(巻貝類;Gastropods)のみ。京大構内でよく見られるのは、所謂でんでんむしやカタツムリやナメクジと聞いてイメージされるものなどの、直腹足亜網(Subclass orthogastropoda)Apogastropoda(下網)異鰓上目(Heterobranchia)有肺類 (Pulmonata)柄眼目(Stylommatophora)の一部。
春夏秋。夜、あるいは雨の時やその前後。吉田南と本部とに挟まれた東一条通。人通りが多い割に気付く人はあまりいないけれども、吉田南に面した石垣を見ると、ほぼ必ず、ナメクジ(Meghimatium bilineatum (種名))が姿を見せています。他の場所にも、総合博物館の石標にはクチベニマイマイ(Euhadra amaliae)、北部構内理学部1号館南東の角付近ではヒラマイマイ(Euhadra eoa)、本部や吉田南の東側の石垣にはキセルガイ(煙管貝: キセルガイ科の総称)、構内のあちこちにはチャコウラナメクジ(Ambigolimax valentianus)、が姿を見せます。北部構内を散策すると運が良ければ毛の生えたでんでんむし、オオケマイマイ(Plectotropis vulgivaga)が見られます。
また、京都大学の生き物マップ – iNaturalist(https://www.inaturalist.org/projects/d2c33586-aefb-4c29-a4fe-0673c8b00912 )によれば、宇治キャンパスではチャイロオトメマイマイ(Trishoplita mesogonia)やウスカワマイマイ(Acusta sieboldtiana)が確認されています。他にも、京都市内の山にはヤマタニシ(Cyclophorus herklotsi)等の有肺類とは別の陸貝も生息していますし、近畿地方だけで約200種の陸貝がいるとされていて、きっと気の付かない所にまだまだいることでしょう。

ヒラマイマイ(北部構内)
今回、かなりの分量となりましたので、目次を置いて措きます:
目次
⚫︎前編:陸貝の身体
・へぇ☆「なめくじとは」
・へぇ☆☆「同じあなで呼吸もうんちもする。」(ツノ/歯舌/雌雄同体/ヤリ/右巻き左巻き/同じ孔/エピフラム/汚れない殻/歩き方/脳)
・へぇ☆☆☆「どうやって歩くのか」
(Direct Wave /もう一つの歩き方)
⚫︎後編:陸貝の行動
・へぇ☆☆☆☆「「アジサイにカタツムリ」は本当なのか。」
・へぇ☆☆☆☆☆「カタツムリの一瞬」
・へぇ☆☆☆☆☆☆「学習し、思考し、葛藤する陸貝。」
・へぇ☆☆☆☆☆☆☆「動物の行動の起源!?」
へぇ☆「なめくじとは」
ナメクジが貝の仲間だと言われて驚かれる方も少なくないと思います。実は、ナメクジのご先祖様は元々、カタツムリのように貝を背負っていたのです。実際、ナメクジ(蛞蝓/ slug/ land slug)とは、殻が退化した陸生巻貝(腹足類)の総称なのです。但し、ナメクジという和名の種(Meghimatium bilineatum;英名: Chinese slug)も有ることに注意が必要です。区別するために「フタスジナメクジ」を和名として用いることもあります。
有肺類では10以上の科レベルの分類群が貝殻を失ってナメクジ化(limacization(limaxがラテン語でナメクジを意味する))しています。サザエの壺焼き等を思い出して頂ければ分かると思いますが、巻貝の内蔵は貝殻内に収められています。ナメクジ化とは内蔵をギュッと頭や腹(足)側に寄せて詰め込むと同時に殻を収縮させて行く過程で、大雑把には次の二段階に分けられます:
段階①: 体表面に外套構造や内蔵の入ったこぶが残っていたりするものの、大雑把に体型が円筒状(ナメクジの形)となっている。
段階②: 体表に内蔵の塊や外套構造の痕跡の一切がない。それらの痕跡は失われているか、頭の少し後ろの腹に近い所の血体腔の内側に収納されてるかである。こぶの無い一様に滑らかな表面をしている。
多くの分類群がナメクジ化して段階①に達している一方、段階②に達しているのは収眼類(Systellommatophora)のみ(海生も含めればmarine doridacean nudibranchs: 裸鰓類(らさいるい: ウミウシの仲間)ドーリス亜目 も)です。しかし、日本の私たちが普段見かけるナメクジであるナメクジ(種名)やチャコウラナメクジは収眼類ではなく柄眼類なのです。つまり、私たちが普段ナメクジと呼んでいるものはナメクジ化の完成形ではないということになります。
チャコウラナメクジは名前に「コウラ」と有る通り、コウラを背負っているのが観察すれば確認できます。ヤマナメクジやナメクジ(フタスジナメクジ)も、殻は完全になくなっているものの、大きな外套膜を背負っています。

チャコウラナメクジ(本部構内)。身体の半分くらいに境目のようなものがありますが、それより前方を覆っているのは外套膜。

ナメクジ(吉田南構内)。お尻の先端だけ色の異なる組織が見えていますが、そこだけ外套膜に覆われていない、それ以外は外套膜に覆われている。
横道に逸れますが、ふつうナメクジやカタツムリと言われると、眼が触覚(ツノ)の先端に位置するという形質を持つものを思い浮かべるでしょう。眼を触覚の先端に持つ有肺類を構成するのが柄眼類と収眼類とで(しかし有肺類の95%は柄眼類([1]))、分子系統解析([5])によると、柄眼類と収眼類で単系統なのではなく、寧ろ触覚の付け根に眼を持つオカミミガイ科が収眼類と単系統を成し、触覚の先端に眼を持つという形質は有肺類の中で二度独立に獲得されたかも知れないとされています。柄眼類と収眼類との違いは、触覚の数と、それを引っ込められるかどうか、です。柄眼類の頭部触覚(ツノ)は大小2対あり、内部には牽引筋があり、触覚を反転させ体内に収容することができますが、柄眼類以外では頭部触覚は1対のみであり反転させることもできません([1])。
本筋に戻りましょう。さて、そもそもどうして陸貝たちはナメクジ化するのでしょうか。
軟体動物(貝類)の殻の存在意義は防御にあるというのが最も一般的な解釈です。硬い殻の中へ閉じこもることで捕食から逃れられるのは容易に想像できます。
また陸に於いて、殻は乾燥からも身を守ってくれます。殻の中に閉じこもってじっと雨を待つカタツムリの姿はよく見かけることでしょう。この乾燥という水中では起こらない陸上特有の状況に対して、海洋で古くから身につけていた殻という形質がたまたま結果的に適応的となっているのです。これは恐らく前適応(preadaptation)の分かりやすい良い例でしょう。
前適応とは、或特定の環境下で適応的な機能を果たしていたりいなかったりする(生物の)形質が、環境の変化等で別の環境に置かれた時にたまたま、その形質のそれまでの環境での適応性とは無関係に、全く別の適応的機能を果たすこと、またはその形質のことです。“形質はそのまま”に環境に対する機能的意味が変化したり多義的になったりする、というものです。前適応の最も古典的で有名な例は鳥の羽です。もともと保温性という機能を持つ形質として獲得された羽毛(それ以前はウロコ)が、いきなり空という環境への適応的機能を果たすように自然選択が働いたとするのには無理があります。ここに進化論への懐疑が生じやすく、翼の獲得には超自然的な力や神の手などが働いたと思わされたり、少なくとも計画的に進化が起こったと思われやすいのです。これに対する進化論の一つの応答が前適応です。つまり、前脚に翼が発達したのは空を滑空したりし始める以前から飛翔とは全く無関係な何らかの適応的機能、例えば抱卵など、を果たすものとしてであり、そうして大きくなった翼の「使いまわし」「転用」にたまたま(抱卵以外の)別の機能の一つとして飛翔が有った、ということです。実際の鳥類(恐竜)の進化は未解明ですが、良い例としてよく用いられてきました。カタツムリの殻も、海洋では単に被食防止という機能の下で適応進化したものだったものが、たまたま陸に於いて乾燥からの避難所になった、前適応と考えられる気がします。
少し横道に逸れてしまいましたが、一般的な解釈に従えば、殻を退化させるというのは、これらの防衛を失うことを意味するのです。しかし実際には、海陸両方に於いて多くの軟体動物が殻を退化させて繁栄しています。殻を持つことには不都合も有るからです。
殻を持つことの主な不都合は次のようなものでしょう:
① 殻を作る費用。炭酸カルシウムを能動的に取り込んで形成するのに掛かるエネルギー。
② 移動の不便。殻の重さはそれだけ移動の際エネルギーを奪う。また殻は移動の障壁となり、行動範囲の制約となる。
③ 浮力の低下。海生の場合、重く丈夫な殻を持つことで海底にしかいられない。浮遊性の軟体動物は、殻自体を浮力調節器官としている頭足類(オオムガイ等)を除いて、殻を薄くしているか持たないかである。
陸貝に当て嵌まるのは①、②です。
特に陸上に於いては殻形成に必要なカルシウムを取り込むのは容易ではありません。海の貝類とカタツムリやマイマイなどの陸の貝類とをイメージして比べてみて下さい。どちらの貝殻が分厚く頑丈で多様性に富んでいるでしょうか。陸貝に薄く似た様な巻き方の殻が多く、多様性に乏しいのは、殻の材料の節約のためです。カタツムリの殻をよく観察すると、一周前の巻きの殻に大部分が被る様に新たな殻が作られて行くのが分かりますが、これは一周前の巻きで作られた部分をできるだけ無駄にせず新たに加える殻を最低限にしようという形態なのです。殻の維持のために雨上がりにコンクリートを齧るカタツムリを見掛けることもしばしばです。コンクリートの主な材料は石灰石ですが、カタツムリも石灰岩の地域に多く生息します。石灰岩自体は、元々、大昔海の底に積もっていた炭酸カルシウムの殻を持つ生き物の遺骸です。カタツムリは悠久の時を跨いだリサイクルをしているのです。因みに、野鳥にとって、カタツムリの殻は貴重なカルシウム源の一つで、特に繁殖期には卵殻の材料として重要となり、産卵数や卵殻の厚さに影響すると言われています。実際、酸性土壌でのカタツムリ減少によるスズメ目の生殖不良の研究があります([6])。

クチベニマイマイ(総合博物館前)殻が薄く透けている。
2019年5月30日午前9時40分、JR九州の設備で停電が発生。列車26本が運休する等の被害がラッシュアワーを直撃し、乗客推定1万2000人に影響が生じました。JR九州が電気設備内部を点検した所、長さ2〜3 cm ほどのナメクジの死骸が発見されました。この小さなナメクジが電気ケーブルに接触、感電して停電が引き起こされたのです。電気設備は虫や小動物が入らないよう接合剤で隙間を固めてられていたのですが、ナメクジは僅かに残る隙間を見逃さず侵入したのです。([7])
ではナメクジは、捕食者や乾燥からの防御を捨てたのでしょうか。いやそうではありません。殻に代わる味方を持っているのです。その一つがあのネバネバヌメヌメネトネトの粘液です。ナメクジはストレスを感じると粘液を大量に分泌します。例えばアリに襲われた時に粘液を絡ませることでアリは身動きを取りにくくなります。また、粘液は、感染症から身を守る免疫系の役割を果たしたり、刺激性の物質を中和させる働きも有るのです。もう一つの武器は身軽さです。粘液はカタツムリも持っていますがこちらはナメクジ特有です。身軽??ナメクジの英語slugの形容詞型sluggishが「のろまな」という意味である通り、ナメクジはのろまなイメージでしょう。それに異論を唱える気はありませんが、カタツムリと比べれば身軽で、僅かな隙間にも入れます。その身軽さを活かして一刻も早く暗く湿った場所へ逃げ込み乾燥や捕食者から逃れるのです。
へぇ☆☆「同じあなで呼吸もうんちもする。」
でんでんむしが愛される理由の一つに、その少しヘンテコな見た目があるのは間違いないでしょう。殻の渦巻き、大きなツノ、ヒゲの様にも見える小さなツノ、大きなツノの先端にちょこんと墨入れしただけのような眼、その他言語化し切れない諸々。ここではそんな陸貝の身体のつくりを解説していきます。
ツノ。ものはほぼ見えない。
先ほどにも書いた様に、私たちがよく目にする陸貝は柄眼類というグループに属していて、頭部触覚と呼ばれる大小2対のツノ(大触覚と小触覚と)を持ちます。童謡の中の「ツノ」は小触覚のみのことだとも言われています。
大触覚の先端には小さな眼があり、一応私たちと同じくカメラ眼(レンズ眼)ではありますが、光の明暗を感じる程度にしか使えないものだと言われています。半径 1〜2 mm くらいの範囲にある小さな物体は認識できるがそれ以外はぼんやりだと言われています([8])。実際、かなり近付いて観察しても気付いたそぶりは殆ど見せません。実は陸貝は夜行性なので重要ではないのです。一方で人には見えない近紫外線を感知する能力をもっているとも言われています。ヒトは波長 400〜700 nm を感知できますが、陸貝は 400 nm より短波長も少し感知できるのです。
また夜行性で乾燥を嫌う陸貝は、左右の大触覚先端の眼で受けた光の強度を比較することでより暗い場所を探しそちらへと移動します。チャコウラナメクジの実験[9]では、片側の大触覚を切除した時(しばらくすると再生する)に切除された側へとぐるぐる旋回する様子が確認されています。切除された方から光が感知されないことでその側の方が暗いと認識して、普段通り暗い方向へ移動しようとする結果、旋回してしまうのです。暫くするとおかしいことに気が付き行動を修正します。
因みに、同じ軟体動物でも、イカやタコといった頭足類では私たちと同様に高精度なカメラ眼を独自に持っています。独自にというのは、軟体動物の眼と脊椎動物の眼とはそれぞれの進化の道筋でそれぞれに獲得されたという意味で、例え似た形質でも系統発生的には両者はほぼ無関係だということです。このように異なる独立な系統発生の先で似たような形質を獲得する進化を収斂進化(しゅうれんしんか)と言います。収斂進化は、軟体動物や無脊椎動物を脊椎動物と比較して見る時にはとても面白い話題です。
小触覚は主に嗅覚や味覚として機能していると考えられています。大小どちらの頭部触覚も触覚(触れた感覚)を持ちます。
陸貝にとって、最も重要な感覚は嗅覚です。触覚はにおいを感知するのみならず、においを記憶する機能も果たしています。チャコウラナメクジの実験[10]では、4本の触覚すべてを切除した後では、その触覚が再生しても、ナメクジは切除前のにおいは全く覚えていないことが確認されています。これはヒトで言うと、自分の眼を一度失って再生できた時に失う前に見た風景を思い出せるか、ということですが、陸貝の場合は(風景でなくにおいですが)これができないのです。つまり嗅いだにおいを触覚で記憶しているのです。またリンゴマイマイの実験[11]では、特定の触覚だけに麻酔を掛けた状態でにおいを覚えさせ、思い出させる時には麻酔を掛ける触覚と掛けない触覚とを逆にした結果、思い出すという行動が示されませんでした。つまり、嗅いだにおいは嗅ぐのに使った触覚で記憶し、嗅いだにおい嗅ぐのに使ったその触覚でしか思い出せない、ということです。
舌はおろしがね、その歯は2万本、食べ跡は幾何学模様。
歯舌(しぜつ)は軟体動物特有の摂餌(せつじ)器官(食事のためのからだの仕組み)です。歯舌は口の中に有る口球(こうきゅう)と呼ばれる舌のような筋肉の塊の上に有り、歯舌膜と呼ばれるリボン状の膜の上に歯舌歯(しぜつし)と呼ばれる小さな歯がおろしがねのように規則的にならんでいます。陸貝の歯舌歯はなんと2万本などといった数に上ります。
ざっくり言えば、表面がおろしがねのようになった舌を持っているのです。陸貝は、例えば岩の表面に着いた緑藻(りょくそう)を、岩の表面をこの舌で舐めるようにして、削り取って食べています。
陸貝の食痕(陸貝が歯舌で緑藻を舐め取った痕跡)はとても幾何学的な美しい模様となります。陸貝の食痕は京大構内でもあちこちで見ることができ、陸貝の姿は見えずともその存在を強く感じることができます。
京大本部構内北門付近の看板に見られる、陸貝の食痕。



こちらの動画も参考にして下さい(流石NHK):陸で生きる巻き貝 カタツムリ | ミクロワールド | NHK for School (https://www2.nhk.or.jp/school/watch/bangumi/?das_id=D0005100141_00000 )
歯舌歯は口腔(こうくう)、つまり口の中ではもちろんむき出しですが、口腔より奥にもむき出しでない歯舌歯は有り、歯舌囊(しぜつのう)とよばれるものに包まれています。歯舌囊の内部には造歯細胞(ぞうしさいぼう)という歯舌歯を分泌、つまり生産するところが有り、擦り減るに従って、その分泌に伴って、前へ前へと新たな歯舌歯が送り出され、前方先端の古い歯舌歯はどんどん切り捨てられ、世代交代していきます。
雌雄同体。二匹で交尾したらどちらも、なんと頭から、卵を産む。
私たちが普段目にする陸貝はすべて有肺類ですが、すべての有肺類は雌雄同体です。
普段私たちが目にする陸貝は、同時的雌雄同体(一つの個体が同時に雄であり雌である)で、性器は必要に応じて生殖孔(せいしょくこう)と呼ばれる頭部の右側の孔(あな)から出され、頭部右側同士を接触させて互いに精子を交換して体内受精します。そして卵(受精卵)もまた、同じ頭部右側の孔(あな)から産み落とされるのです。但し、ここで「右側」と言った部分はキセルガイ科やヒダリマキマイマイ等一部の種類や、右側の種でも変異個体では、「左側」になります。
「ヤリ」って何?
「ツノ出せヤリ出せ目玉出せ~♪」。有名な歌詞にある「ヤリ」、これは一説によると、恋矢(れんし;love dart)と呼ばれる器官のことを指しているのではないか、と言います。
恋矢は、石灰質で硬く鋭利で刀剣のような器官で、どのように使うかと言うと、なんと交尾の時に身体から出して相手に突き刺すのです。恋矢は、相手の体に刺さると、恋矢表面の粘液が相手の体内に注入されます。この粘液は、相手に受け渡した精子が相手に分解されることや、相手が別のカタツムリと再び交尾をすることを、防いだりする働きがあります。なので、喩えるなら、自分の精子をより多く相手の卵子と受精させるために恋矢を刺すのです。しかし一方で、刺した交尾相手の寿命を3/4にまで縮めてしまい、さらに交尾相手の生涯の産卵数を減らしてしまうことも分かっています([12])。相手を傷つけてでも自らの子孫繁栄に有利な行動を取る、「利己的な遺伝子」の仕業なのでしょう。
しかし、この「ヤリ=恋矢」説、あの「ダーウィンが来た」 「知らなかった!カタツムリ大研究」 – ダーウィンが来た! – NHK でも推されていましたが、実はかなり疑わしい説なようで、実際は大触覚のことを指しているのではないか、という考えもあり、筆者的にもその方が自然に思います。では「ツノ」は?となりますが、そう、「ツノ」は小触覚となります。大小どちらが「ツノ」でどちらが「ヤリ」かももしかしたら違うかも知れません(歌詞の順番的に大触覚が「ヤリ」という方が自然に思えます)。そもそもこの童謡は昔各地に伝わっていたわらべうたを起源とするもので、何が「ヤリ」で何が「ツノ」か統一された定義は無かったかも知れません。
(参考:[13])
右巻きまれに左巻き
陸貝は巻貝に含まれる生き物です。殻を持つ巻貝には殻を巻く向きが有り、大きく、右巻きと左巻きとが有ります。上から(殻頂方向から)見た時、時計回りに拡大する螺旋なら右巻き、逆巻きなら左巻きです。他の見分け方としては、殻を縦に置いて殻の口の正面の側から見た時、殻の口が右側に見えれば右巻き、左側に見えれば左巻きです。


右巻き(上(殻頂方向)から見て時計回りの付加成長)
私たちがカタツムリとして良く知る種の貝殻は一部例外を除いてほぼすべて右巻きです。
先程、性器を出したり卵を産んだりする孔(あな)も一部例外を除いて頭部右側と言いましたが、実は、この右というのは右巻きという巻きの向きに対応しています。孔(あな)が頭部右側にあることと右巻きであることは同義で、孔(あな)が頭部左側にあることと左巻きであることは同義なのです。このことは種内変異でも同じで、巻きの左右と孔の左右は対応します。
このことが何を意味するかというと、交尾が右巻き同士か左巻き同士かでしかできないのです([14])。なので、例え左巻きの変異個体が現れても、同じく左巻きの変異個体と出逢わない限りその子孫は残らず、基本的には、右巻きの種は右巻きの個体しか増えないのです。例外はキセルガイ科で、キセルガイ科の交尾は、殻が細長く丸っこいものよりも邪魔になり難く、一方が他方の上に乗るような姿勢が取れて右巻きと左巻きとでも一往成立するのです。殻が細長い分類群ほど左巻きの種が顕著に多いことも知られています。約200種とされる日本産のキセルガイ科は全種左巻きです。右巻きのキセルガイが見つかっただけでニュースにもなりました([15])。
同じ巻の向き同士でしか交尾できないにも関わらず左巻きの個体も少なくない、という例外もあります。これには「右利きのヘビ仮説」というとても面白い研究が有ります([16],[17])。この仮説は、とても端折ってざっくり言えば、右巻きのカタツムリを捕食するのに特化して「右利き」という行動様式を獲得したヘビの影響により、左巻きの個体がヘビに食べられ難く数を残し易くなったことで、左巻きの個体が比較的多く存在するというものです。実際にヘビはどうやってカタツムリを捕食するのか、どう右利きなのか、詳しいことは参考文献[16],[17]をご覧下さい、特に[16]は仮説提唱者である細将貴氏が論文[17]執筆に至る物語のようなもので、読み易く面白いと思います。私たちが目にするカタツムリはキセルガイを除いて殆どが右巻きですが、それは或意味、「右利きのヘビ」の不在ということなのです。
左巻き右巻きの話は未だに多くの謎を残していて、様々に研究されています。また何かおもろい話が飛び出すことに期待が高まります。

日本産キセルガイの全種が左巻き。(写真は吉田南)
同じあなで呼吸もうんちもする。
陸貝には私たちでいうところの鼻の孔のような部分は有りません(臭いは触覚で嗅ぐのでした)し、口で呼吸はしません。私たちがよく知る陸貝は肺呼吸(有肺類)ですが、呼吸は呼吸孔(こきゅうこう)というそれ専用の孔を通して行われます。この呼吸孔、その出口をなんと排泄孔(うんちを出す孔)の出口とほぼ同じくしているのです。「え~….」と不快感を覚える方もおられるかも知れませんが、恐らくその不快感は、私たちにとって呼吸といえば、匂いや味を感じたり食事したり食道へ直接通じていたりする器官である、鼻口でするもの、であるからでしょう。ですが陸貝の呼吸孔は食道に直接通じていたりしませんしそこで匂いや味を感じることはありませんので、そこまでありえないものでもありません。おまけに、基本、陸貝の糞は臭くありません。
これらの孔(あな)、カタツムリだと身体を殻から出して活動している時は殻の口(殻口)付近(これは身体の右側)に見ることができ、ナメクジでも身体の右側の生殖孔のすぐ後ろに見られます。呼吸孔、排泄孔も右巻きと左巻きに対応しているのです。
エピフラム(epiphragm)
実は陸貝(特に私たちの身の回りの種は)、冬眠はもちろん、夏眠もします。変温動物である陸貝は、寒いと動けなくなる上に、暑さにも弱いのです。乾燥に弱いため、乾燥した時も休眠します。カタツムリの場合、休眠中乾燥や天敵からの捕食を防ぐために殻の口に蓋をします。この蓋は普段どこにも有りませんが、休眠する時に自ら作るのです。この蓋をエピフラム(epiphragm)と言います。
種によってはカルシウムの硬い蓋を作ることも有りますが、エピフラムは多く場合、粘液で作られ、それが乾いて固まったものです。休眠中のカタツムリが葉っぱの裏などに殻の口でくっついていることもありますが、これも粘液で作られたエピフラムが接着剤となっているのです。成長しきった大人のカタツムリの殻の口は外にそり返っていて、この様にピタッと殻の口をどこかに付けて乾燥を防ぎ易い機能を果たすと考えられています。乾燥が強かったりすると、カタツムリは殻の奥へ奥へと引っ込もる中で幾度かエピフラムを作り、幾層かの蓋がある構造となることもあります(オオムガイの隔壁みたい…)。

休眠中のクチベニマイマ(総合博物館前)。
汚れない殻。フラクタル。温暖化で色が変わる!?
カタツムリは、陸上への適応として、「汚れない殻」を獲得しました。
カタツムリの殻は硬タンパクのみからなる殻皮層と、薄いタンパク質のマトリックスで囲まれたアラゴナイト結晶からなる石灰質層3層(稜柱層、層板層、真珠層)とで、層を成しています。この殻皮層というタンパク質の層が、濡れにくさや汚れにくさの要因だと言われています。
また、カタツムリの殻表面の硬タンパク質層には、約 0.5 mm 間隔の成長線の(殻口から新たな殻を付加して成長していく過程でできる)溝と、それに直交する約 0.1 mm 間隔の規則正しい溝が形成されている上、雁行状(雁(ガン)が群を成して飛ぶ時の様に斜めの線状)のしわが約 10 μm 間隔で並んでいます。これがフラクタル構造となっていて、フラクタル次元は2.1とさほど歪(いびつ)ではないものの、その次元が 0.1 μm から 800 μm という広いスケール間で維持されていて、フラクタル構造の範囲を広げることで、微小範囲での大きな表面積を獲得し、更に汚れを弾きやすくしているのです。

格子状に細かく溝が走るクチベニマイマイの殻。
特に殻を水に濡らしている時に、油を強力に弾くのだとか。
実際どれほど汚れにくいのか。殻に油性ペンで何か書いてみた後、水で洗ってみるとしつこいはずの油性ペンの汚れが簡単に流されるのです。実際に試してみるといいかも知れません。
陸上進出という進化の一大イベントの中でこの性質を獲得したと言われていて、人工物への応用も様々検討されています([18],[19])。
そんなカタツムリの殻ですが、模様や色彩も多様性に富んでいます。その多彩さは、ハワイ出身の宣教師ジョン・ギュリック(John Thomas Gulick)を魅了し、彼はカタツムリの殻の多様性から様々な着想を得てダーウィンが創始した直後の進化論を発展させ時代の先を行く理論を作り、日本の貝類学発展のきっかけも作りました([20])。(一方でハワイのカタツムリの幾種かはギュリックが採り過ぎたことが要因で絶滅したとも言われていたりします。)しかしそんな殻の色彩の多様さも地球温暖化の中で失われてしまうかも知れません。温暖化が進んだ環境を模した実験[21]では、色の濃い殻を持つカタツムリの熱耐性が低く、体温上昇と水分不足が原因で色素の強いカタツムリほど死亡率が高いことが示されています。結果として色素の弱い白っぽい殻のカタツムリしか生存しないような「熱選択圧(thermal selection pressure)」が、カタツムリの進化等に影響を及ぼすというのです。
脳で直接明るさを感じる。
陸貝の脳?驚く方も多いかも知れませんが、カタツムリやナメクジなど、陸貝にも脳が有るのです。しかしこの脳も収斂進化の産物です。「脳」は狭義には脊椎動物の脳を指す言葉ですが、無脊椎動物でも機能や形態などの類似から「脳」と呼ばれる器官を持つのです。脊椎動物とは独立した系統発生を辿ったからこそのパラレルワールド的魅力が有ります。
私たちのよく知る陸貝の脳は大小四つの触覚を根元へ延長してだいたい交差する位置にあります。そしてなんとその脳のど真ん中を食道が突っ切っているのですが、実はこれは無脊椎動物ではメジャーなボディープランです。脳を構成するニューロン(神経細胞)の数はチャコウラナメクジでは数十万個とヒト(数百億個)の10万分の1程度と、やはりヒトや哺乳類の脳より単純とも言えそうなものですが、損傷しても再生する能力を持っていたりのヒトに無い能力を備えています。
特に驚くべきは、脳が光を感知できるということです。チャコウラナメクジの実験[23]によると、眼の着いた大触覚を切除した盲目な筈のナメクジが明かりに晒されると、切除されてないナメクジに比べて紆余曲折しつつも、最終的に暗い場所を探し当てるのです。ナメクジの脳には光(特に400〜480nmの短波長)に反応する特殊なタンパク質があり、光の方向等は感知できないけれども、その瞬間感知している光の強度と直前のそれとを比較してより暗い方へと移動することが出来ると考えられています。参考文献[23]のurlでは実験で観察されるナメクジの行動を動画で見ることができます。カタツムリも脳で光を感知すると考えられています([8])。
へぇ☆☆☆「どうやって歩くのか」
私たちは陸貝の移動の仕方を「這う(はう)」と簡単に表現しますが、では「這う」とはどうしていることなのか、と訊かれて説明できる人はどれほどいるのでしょう。陸貝の「這う」は、ヒトが地面を這う仕方とは全く異なりますが、同じ語であるが故に少々混同して、陸貝は不器用に身体を無理矢理引き摺るように移動しているとイメージしている節があるのではないでしょうか。しかし実は物理現象をたくみに利用した驚きの方法で優雅に移動しているのです。
Direct Wave
巻貝は「腹足類」という名の通り、腹足とも呼ばれる軟体部と地面が接する「腹」全体を「足」として運動させることで移動します。これが「這う」こと、這行運動(しゃこううんどう; crawling locomotion, adhesive locomotion)です。巻貝が透明な板の上を「這う」様子を裏から見ると弛緩収縮(しかんしゅうしゅく)の模様(筋収縮パターン)を見ることができます。巻貝の移動に伴ってこの筋収縮パターンは波打ちます。ふつう、陸貝の場合はこの波の進行方向が陸貝自身の進行方向と一致します。海の貝では別の色々な移動の仕方があり、
下から見てみる | カガクノミカタ | NHK for School
(https://www2.nhk.or.jp/school/watch/bangumi/?das_id=D0005110365_00000 )
を是非ご覧下さい(流石NHK)。この動画の中で紹介されているシッタカガイやサザエの様に左右非対称な筋収縮波を ditaxic wave 、陸貝(e.g. カタツムリ)の様に左右相称な筋収縮波を monotaxic wave と言い、左右相称(monotaxic wave)でもヒザラガイの様に身体の進行方向と逆向きならば retrograde wave 、陸貝(e.g. カタツムリ)の様に同方向ならば direct wave または prograde wave と言います。
筋収縮波は、伸長と収縮とが交互に配置されたパターンになりますが、direct wave と retrograde wave の違いは至極単純です。収縮している部分を自由に動くようにし、伸長部分を地面に固定して動かさない、この条件だけで筋収縮波は direct wave となります。逆に、収縮している部分を地面に固定し動かさず、伸長部分を自由に動くようにする、この条件だけで筋収縮波は retrograde wave となります。つまり、direct wave と retrograde wave の違いは、収縮と伸長のどちらを自由に動かし、どちらを地面に固定して動かさないかの違いなのです。
私たち等、多くの陸上脊椎動物は retrograde wave を生んで運動しているとも或意味言えます。というのも、歩く時、歩幅を生んで足で持ち上げられた(浮かせた)身体を伸長させて運動していますよね。上の動画のヒザラガイを見ても、retrograde wave はとてもイメージし易いかと思います。一方の direct wave は、何かがそれで移動している様子を見かけることは(ヤスデなどが歩く様子はそうですが)殆どなく、とてもイメージが難しいと思います。
なので、手書きですが、図を作ってみました:
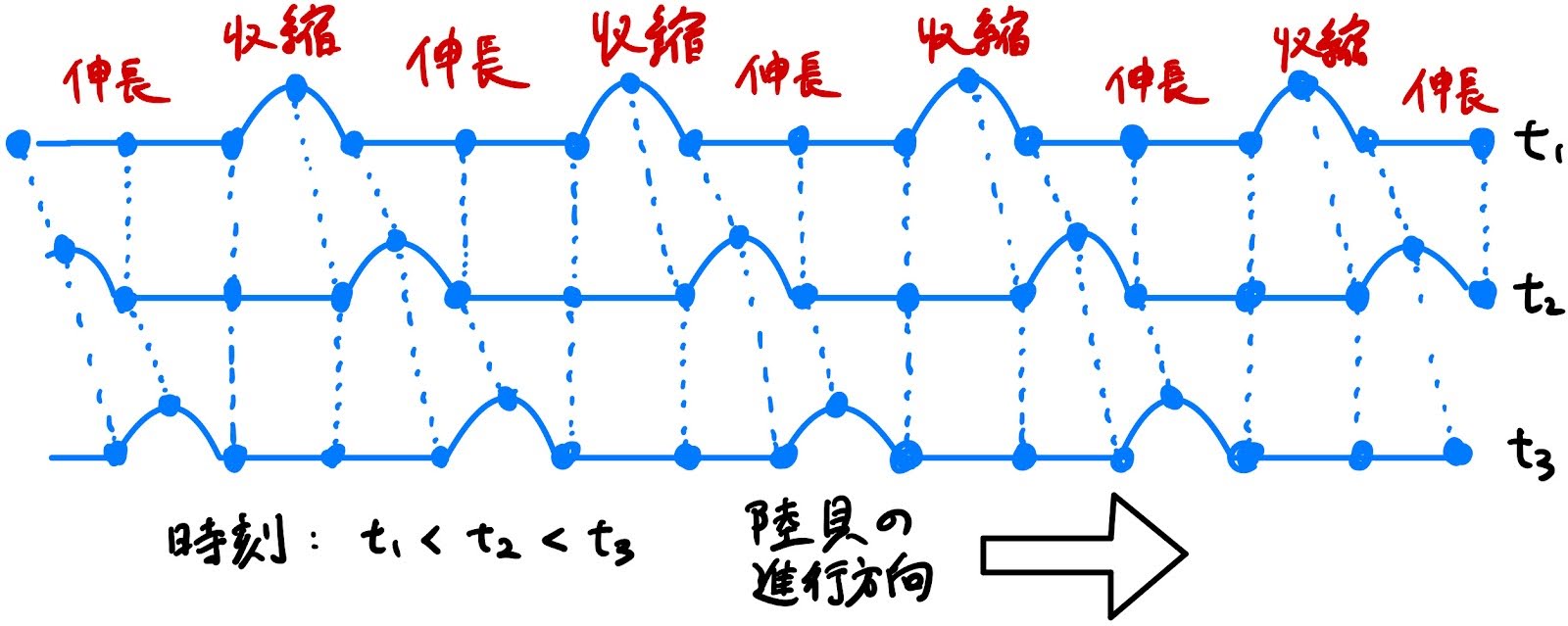
この図は陸貝というよりヤスデの歩き方の図と言った方が誤解がない。しかし本質的には同じことで、伸長部を地面に摩擦により固定し、一方で収縮部を(滑らせるか浮かせるかして)摩擦から解放することで、direct wave となる、前進する。(手書きの図ですみません…)
この図で先ず分かることは、伸長した部分は接地させて(図の直線部分)収縮させた部分を浮かせる(図の曲線部分)、という約束の下で、伸長-収縮の波(筋収縮波)を生み出せば、その波の伝わるのと同じ向きへと全体が進行する、ということです。これはより一般には、収縮部分で地面との摩擦を小さくし、伸長部分で地面との摩擦を大きくする、という摩擦制御により direct wave となる重心移動が可能になるということです。収縮部分を持ち上げ地面から離す、というのは、この摩擦制御の特殊な一例に過ぎません。
余談になりますが、収縮部分を持ち上げるという方法での摩擦制御による direct wave で歩く生き物がヤスデです。
ヤスデの歩きはこちらの動画の2’55″〜みれます(流石NHK):
いきものの なみ | ミミクリーズ | NHK for School( https://www2.nhk.or.jp/school/watch/bangumi/?das_id=D0005260179_00000 )
本題に戻って陸貝の direct wave ですが、地面と接する部分と接しない部分とを使い分けるような方法とは、どうやら別の方法で摩擦制御をしているようです。鍵を握るのが粘液です。陸貝の這行運動に粘液が関係すると、具体的な実験結果を根拠にして最初に示唆したのは、アメンボ水面走行に関するDennyの逆説(アメンボの記事で紹介しました:https://eco.kyoto-u.ac.jp/?p=9161 )の、あのDennyさんです。Dennyはバナナナメクジ(Ariolimax columbianus )を用いた実験[25]で、粘液は力の掛かり方に応じて、弾性固体(ぷにぷに?)の性質を示したり粘性流体(どろどろ?)の性質を示したりする、つまり、粘液の性質は力の掛かり方で固体的か液体的かがスイッチする、ということを発見し、これが這行に関係するのではないかと考えたのです。その後の研究や数理モデルなどで具体的にどう関係するのかが明らかになってきました。それによると、腹足と地面間にある粘液は何の動きも無い時は弾性固体ですが、筋肉収縮により粘液に力が掛かりその力が或大きさを超えたら粘液は素早く粘性流体に変化し、続いて収縮波が去る際に粘液に掛かる力が或大きさを下回ったら粘液は弾性固体に戻る、これにより、普段は弾性固体として地面との摩擦を生んで滑らないようにしている粘液が、筋収縮波の収縮部分下では瞬間的に粘性流体となり液体的に振る舞うことで摩擦を減じるという摩擦制御を示すのです。収縮部分で地面との摩擦を小さくし、伸長部分で地面との摩擦を大きくする、という direct wave による重心移動の条件がこうして満たされているのです。陸貝はこうして全く身体を浮かせずとも歩くことができる、小さな筋力だけで歩けるのです。
粘液に備わった物理学的な性質を用いることで、とても単純に、筋収縮波を伝えるというだけで、とても小さな筋力で、陸貝の移動が可能となるのです。
もう一つの歩き方
陸貝の歩き方には這行運動以外にもう一種類あります。陸貝は状況に応じて二種類の歩き方を使い分けるのです。もう一つの歩き方とは loping gait と呼ばれるものです。筆者は未だ見たことがないのですが、カタツムリがまるでヒトが歩いたような足跡、粘液に濡れた部分が点線の様に間隔を空けてポツポツと続く様な足跡を残すのを、或はその足跡を、目撃した方もおられるのではないでしょうか。この足跡こそ、loping gait によるものなのです。どんな歩き方か幾つかのYouTube動画のurlを載せておきます:
・https://youtu.be/x3g0Ee7cS68 (足跡が分かりやすい)
しかしこの陸貝たちもいつもこうした歩き方(loping gait)をしているのではありません。では時々この歩き方をするのはなぜかというと、諸説ありますが、乾燥して吸水性のある所(乾燥したコンクリートや木材の上)を歩く時に loping gait をすること([26])から、出来る限り接地を減らして粘液の消費を削減しようとしている、というのが恐らく最も有力な説です。のんびりゆったりしているような陸貝たちですが、そうまでするとは、生きるために案外必死に歩いているのでしょうか…。
いかがでしたか??
私たちは生き物の形を見るとき、ついつい自分たちヒトの身体になぞらえて言語化して見てしまいます。しかし陸貝の身体はそのような見方では上手くいきません。陸貝の身体のことを知って、驚いたり、困惑したりしたのではないでしょうか。ヒトの身体でなぞらえようとすると困惑するのです、知らんけど…。ですが、似ていない所ばかりではなく、似ている所があることも分かって頂けたかと思います。でもそれも、(その部分が同じ呼び方をされているとしても)似ているに過ぎず、系統発生的には無縁なのです。
陸貝の身体は、似ていないか、ただ似ているだけ。
後編では、そんな身体を持つ陸貝がどんな行動をするか、行動観察から学習理論を通して見える「陸貝の学習」等を中心に紹介します。
因みに、渦巻の殻と言えばアンモナイトを思い浮かべる人も多いでしょう。アンモナイトはイカやタコの仲間で、イカやタコも元々アンモナイトと同じように身体の外側に殻を持っていたとされています。つまりイカやタコも広い意味では「ナメクジ化」した生物だと言えます。また、アンモナイトは基本的には左右相称ですが、異常巻アンモナイトと呼ばれるヘンテコな巻き方の殻を持つアンモナイトの中には、左右非相称で、右巻きと左巻きの個体があるものが知られています。因みに異常巻陸貝も存在します([27])。
アンモナイトが生きていた時代のような大昔の、アンモナイトを含む炭酸カルシウムの殻を持っていたような生物の、遺骸が海底に長い年月を掛けて積もり、更に長い年月を掛けた地殻変動で石灰岩として地上の岩となり、そこに棲むカタツムリの殻となったり、更にコンクリートとなってから街中のカタツムリの殻となったり…。カタツムリの殻とアンモナイトの殻は似ているだけでなく、もしかすると、あなたの足下にいるそのカタツムリの殻には、大昔アンモナイトの殻だった炭酸カルシウムが含まれているかも知れません。
(筆者の卒業研究の対象はアンモナイトです。)
参考文献
[1] 佐々木猛智『貝類学』東京大学出版,2010.
[2] 松尾亮太『考えるナメクジ』さくら舎,2020.
[3] 野島智司『カタツムリの謎 日本になんと800種! コンクリートをかじって栄養補給!?』誠文堂新光社,2015.
[4] Simone, Luiz. (2018). Malacopedia Main processes of body modification in gastropods: the limacization. 1. 12-22.
[5] Schrödl, M. (2014). Time to say “Bye-bye Pulmonata”? Spixiana 37:161–164.
[6] Graveland, J., van der Wal, R., van Balen, J. et al. Poor reproduction in forest passerines from decline of snail abundance on acidified soils. Nature 368, 446–448 (1994). https://doi.org/10.1038/368446a0
[7] 「JR停電「犯人」はナメクジ 電力設備に侵入、ショート」西日本新聞 2019/6/22 https://www.nishinippon.co.jp/item/n/520710/ (最終閲覧2024/9/19)
[8] RICHARD M. EAKIN, JEAN L. BRANDENBURGER, Understanding A Snail’s Eye at a Snail’s Pace, American Zoologist, Volume 15, Issue 4, November 1975, Pages 851–863, https://doi.org/10.1093/icb/15.4.851
[9] Matsuo, Y., Uozumi, N. and Matsuo, R. (2014). Photo-tropotaxis based on projection through the cerebral commissure in the terrestrial slug Limax. J. Comp. Physiol. A 200, 1023-1032. https://doi.org/10.1007/s00359-014-0954-7
[10] Yurika Koga, Yuko Matsuo, Ryota Matsuo “Olfactory Memory Storage and/or Retrieval Requires the Presence of the Exact Tentacle Used During Memory Acquisition in the Terrestrial Slug Limax,” Zoological Science, 33(1), 78-82, (1 February 2016)
https://doi.org/10.2108/zs150128
[11] Friedrich, A., Teyke, T. Identification of stimuli and input pathways mediating food-attraction conditioning in the snail, Helix. J Comp Physiol A 183, 247–254 (1998). https://doi.org/10.1007/s003590050252
[12] Kimura K & Chiba S (2015) The direct cost of traumatic secretion transfer in hermaphroditic land snails: individuals stabbed with a love dart decrease lifetime fecundity. Proceedings of the Royal. Society London B, DOI: 10.1098/rspb.2014.3063.
[13] 童謡『かたつむり』の「ヤリ」を恋矢だと言うのはもうやめて – note https://note.com/maimaikeikaku/n/n22c6d6c98635?magazine_key=m99db09119e7f
[14] Asami T, Cowie RH, Ohbayashi K. Evolution of mirror images by sexually asymmetric mating behavior in hermaphroditic snails. Am Nat. 1998 Aug;152(2):225-36. doi: 10.1086/286163. PMID: 18811387.
[15] 「逆巻きのキセルガイ発見、京都/突然変異」四国新聞,2010/4/27 11:46
http://www.shikoku-np.co.jp/national/science_environmental/20100427000175 (最終閲覧2024/10/5)
[16] 細将貴『右利きのヘビ仮説 追うヘビ、逃げるカタツムリの右と左の共進化』東海大学出版部,2012.
[17] Hoso, M., Kameda, Y., Wu, SP. et al. A speciation gene for left–right reversal in snails results in anti-predator adaptation. Nat Commun 1, 133 (2010). https://doi.org/10.1038/ncomms1133
[18] 井須 紀文, カタツムリに学ぶ表面防汚技術, 溶接学会誌, 2009, 78 巻, 3 号, p. 187-190, 公開日 2014/02/12, Online ISSN 1883-7204, Print ISSN 0021-4787, https://doi.org/10.2207/jjws.78.187, https://www.jstage.jst.go.jp/article/jjws/78/3/78_187/_article/-char/ja
[19] 井須 紀文, カタツムリに学ぶ住まいの防汚技術, 応用物理, 2013, 82 巻, 5 号, p. 426-428, 公開日 2019/09/27, Online ISSN 2188-2290, Print ISSN 0369-8009, https://doi.org/10.11470/oubutsu.82.5_426, https://www.jstage.jst.go.jp/article/oubutsu/82/5/82_426/_article/-char/ja
[20] 千葉聡『歌うカタツムリ: 進化とらせんの物語』岩波書店, 2017, ISBN: 4000296620, 9784000296625, 202 pages.
[21] Köhler H-R, Capowiez Y, Mazzia C, et al. Experimental simulation of environmental warming selects against pigmented morphs of land snails. Ecol Evol. 2021; 11: 1111–1130. https://doi.org/10.1002/ece3.7002
[22] Kathryn Knight; Slugs ‘see’ with light-sensitive brain. J Exp Biol 15 July 2019; 222 (14): jeb209908. doi: https://doi.org/10.1242/jeb.209908
[23] Haruka Nishiyama, Akane Nagata, Yuko Matsuo, Ryota Matsuo; Light avoidance by a non-ocular photosensing system in the terrestrial slug Limax valentianus. J Exp Biol 15 July 2019; 222 (14): jeb208595. doi: https://doi.org/10.1242/jeb.208595
[24] 岩本 真裕子, 腹足類の這行運動に見る運動メカニズムと制御, 応用数理, 2016, 26 巻, 2 号, p. 14-21, 公開日 2016/09/30, Online ISSN 2432-1982, https://doi.org/10.11540/bjsiam.26.2_14, https://www.jstage.jst.go.jp/article/bjsiam/26/2/26_14/_article/-char/ja
[25] Denny, M. The role of gastropod pedal mucus in locomotion. Nature 285, 160–161 (1980). https://doi.org/10.1038/285160a0
[26] McKee A, Voltzow J, Pernet B. Substrate attributes determine gait in a terrestrial gastropod. Biol Bull. 2013 Feb;224(1):53-61. doi: 10.1086/BBLv224n1p53. https://www.journals.uchicago.edu/doi/10.1086/BBLv224n1p53
[27] Reuben Clements, Thor-Seng Liew, Jaap Jan Vermeulen and Menno Schilthuizen, Further twists in gastropod shell evolution, Biology Letters, volume 4, number 2, pp.179-182, 2008, doi: 10.1098/rsbl.2007.0602
————–以下お知らせ————–
へえ☆4つ以上の情報をご存じの方は、是非SNSでコメントの投稿をお願いします。みなさんの「へぇ」知識の共有をお待ちしております!
また、京大!バイオスクープでは、京大内にあるいきものの疑問や気になることの調査依頼をお待ちしております。こちらも同様にご連絡ください。SNSのコメントやこちらにお送りください。
